
【技术交流】生态修复|东北山地山口湖生态系统的营养结构和演变趋势
东北山地山口湖生态系统的营养结构和演变趋势
石展耀,张靖天*,黄炜惠,翁南燕,张含笑,霍守亮
环境基准与风险评估国家重点实验室,中国环境科学研究院
摘要:为探究东北山地湖泊山口湖生态系统的食物网结构并预测更合理的生态管理方式,结合多元逐步回归分析探索了理化因子对山口湖初级生产力的影响,使用Ecopath模型对2014年山口湖生态系统数据进行建模,并利用Ecosim模型分析不同情景下浮游生物和主要鱼类自2014年开始未来20年的变化趋势,结合相关性分析探究山口湖未来的管理方式。结果表明:山口湖是磷限制型湖泊,水温和总磷对初级生产力的增加有促进作用。山口湖生态系统生物之间捕食关系复杂,能量流动集中在第Ⅱ营养级以上,关键种为“其他鱼类”功能组,山口湖Ecopath模型的Pedigree指数为0.537,可信度较高。浮游植物的生产率(PD/B)增加显著促进了鲫、鲤和鲢相对生物量的增加,PD/B每年下降超过5%时对上述3种鱼类相对生物量的影响不显著,鲢搜索率的增加会提高鲢对桡足类、枝角类和轮虫等浮游动物的捕食效率,通过营养级间联合作用导致浮游植物的相对生物量略微上升。结合情景分析和相关性分析发现,增加浮游植物的生物量会提高渔业产量,山口湖生态系统中鲢控藻效果不佳,要加强对外源营养盐的限制。
关键词:Ecosim模型 / 营养结构 / 生态系统演变 / 山地湖泊
渔业经济的发展会对湖泊生态系统健康状态产生重要影响,过度捕捞特定鱼种[1]、增加投饵投料[2]等渔业活动均会导致水生态系统结构和功能退化。探索水生态系统中生物之间的相互作用对理解渔业活动和生态系统之间的平衡关系,揭示影响湖泊生态系统健康状态的因素具有重要意义。现有研究大多集中于探究入湖营养物、沉积物污染等对湖泊生态环境的影响[3-5],对生态系统中各生物之间关系的探究较少。
EwE(EcopathwithEcosim)模型是一种常用的研究食物网内物质和能量平衡的复合模型,包含Ecopath和Ecosim等模型,该模型基于营养动力学原理和能量守恒方程,能够了解生态系统内各种生物的营养结构组成和能量流动效率,揭示不同生物对生态系统产生的上行效应和下行效应[6],为渔业经济和水生态健康之间的平衡提出发展建议。EwE模型于1999年引入中国并应用于太湖[7]、千岛湖[8]、洱海[9]、洪泽湖[10]、白洋淀[11]等地,用于探讨生态系统的营养结构和成熟度等方面,EwE模型应用的结果表明:渔业经济发展使得生态系统内能量流向主要养殖鱼类,而其他鱼类和浮游动物的生物量呈下降趋势,从而影响生态系统的稳定性和恢复力。Lassalle等[12]构造的比斯开湾的Ecopath食物网分析证明了渔业养殖导致19.3%~39%的能量流向碎屑。邓悦等[13]利用Ecopath模型评价鲢、鳙放养对千岛湖生态系统影响时发现,低营养级能流在生态系统中占比较大,越往顶级占比越小,随着鲢、鳙养殖规模的扩大,2004—2016年流向碎屑的能量占比由53.3%增至64.3%。现有的EwE模型研究大多集中于中东部湖泊渔业经济发展对生态系统的影响,关于高寒山地湖泊生态系统结构和演替趋势的研究较少。
东北平原山地湖区地处温带湿润、半湿润气候区,夏季降雨丰沛、冬季积雪不化,年均气温在5℃左右,污染物降解速度慢,每年长达4~6个月的冰封期阻断了外源营养物的输入,降低了生态系统的物质能量循环效率,提高了水体的富营养化水平[14]。低温导致藻类、浮游动物和鱼类等生物生长缓慢,微生物降解速度降低,进一步弱化了水体的自净作用。因此,研究高寒山地湖泊生态系统的营养结构和能量循环可以为水体污染控制和渔业经济发展提供支撑。笔者选择具有典型东北山地湖泊特征的山口湖[15]为研究对象,采用EwE模型探索山口湖生态系统的营养结构、能量传递效率并确定其关键种,利用Ecosim进行情景分析预测山口湖中浮游生物和主要鱼类之间的关系并预测发展趋势,以期为山口湖渔业和生态系统平衡可持续发展提出可供参考的建议。
1. 材料与方法
1.1 样品采集与分析
山口湖位于嫩江一级支流的讷谟尔河上游,地处小兴安岭西坡余脉向松嫩平原延伸的过渡地带,湖区周边森林相对密集,低山、丘陵及开阔U型河谷内沼泽和湿地发育。山口湖水域面积84km2,平均水深16m。总库容9.95亿m3,平均年入湖总径流8.2亿m3。山口湖始建于1995年,1999年竣工验收,目前是以防洪、发电、供水、生态用水、旅游并举,兼顾灌溉、水产养殖等综合利用的水利工程,湖内常年养殖鲢、鳙。
山口湖由南北河、长水河、土鲁木河等河流汇集后建坝形成。2014年,在山口湖开展了14个点位(图1)的水质调查工作,采样时间为2014年1—3月、5—10月、12月(4月和11月由于水冰交融无法采样),分析水温(T)、化学需氧量(CODCr)及溶解氧(DO)、总氮(TN)、总磷(TP)、叶绿素a(Chla)浓度;分别于2014年6月、7月、10月、12月开展了4次山口湖浮游植物、2次浮游动物、底栖动物调查工作;同时,对2014年山口湖渔业产量和组成进行了资料调研。
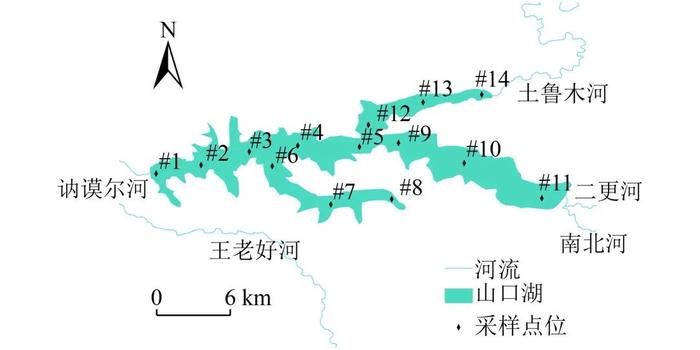
图 1 山口湖采样点位分布
Figure 1. SamplingsitesinLakeShankou
1.2 相关性分析
采用Spearman相关性分析筛选出影响生态系统初级生产力的重要因子,即T、DO、CODCr、TP、N∶P(质量比,下同),利用多元逐步回归模型对Chla进行分析,根据其中PF(即各项因子F检验得到的P)大小依次向方程中引入变量T、CODCr、DO和TP,从数值上确定环境因子对山口湖初级生产力的响应关系。
1.3 Ecopath和Ecosim模型原理
Ecopath模型以2个主要的生物质量方程为基础:式(1)用于描述功能组生产量在生态系统内的流向;式(2)用于描述各功能组的能量守恒[16]。
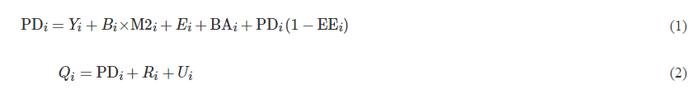
式中:PDi为功能组i的生产量,t/(km2·a);Yi为渔业捕捞量,t/(km2·a);M2i为被捕食死亡率,a−1;Bi为生物量,t/km2;Ei为净迁出生物量,t/(km2·a);BAi为累积生物量,t/(km2·a);EEi为功能组i的生态营养效率;Qi为总消费量,t/(km2·a);Ri为总呼吸量,t/(km2·a);Ui为未被利用的能量,t/(km2·a)。
Ecosim模型反映了功能组生物量在一定时间范围内的变化,其由Ecopath模型的方程求导后得到的动力学微分方程:
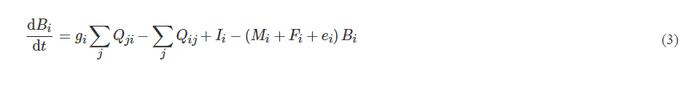
式中:dBi/dt为功能组i的生长速率,t/(km2·a);gi为净生长效率;Mi为非捕食的自然死亡率,a−1;Fi为捕捞死亡率,a−1;ei为净迁出率,a−1;Ii为生物量净迁入率,t/(km2·a);
Qji为功能组j被功能组i所捕食的效率,t/(km2·a);Qij为功能组i被功能组j捕食的效率,t/(km2·a)。
1.4 山口湖EwE模型的应用
功能组的分类。参考李昌等[17-18]的研究成果,根据2014年山口湖的采样调研数据和不同物种的生态位将山口湖的生物分为17个功能组:其中包括鲢、鲤、鲫、草鱼、狗鱼、哲罗鱼、密苏里白鲑、鲶和其他鱼类9个鱼类功能组(其他鱼类功能组包括鳌花、鳊花、鲤花、哲罗、法罗、雅罗、同罗和胡罗等低生物量鱼类),浮游植物、桡足类、枝角类和轮虫4个浮游生物功能组,以及水生植物、原生动物、底栖动物和碎屑4个功能组,功能组分类见表1。
表 1 山口湖Ecopath模型功能组输入与输出参数
Table 1. InputandoutputparametersoftheEcopathmodelinLakeShankou
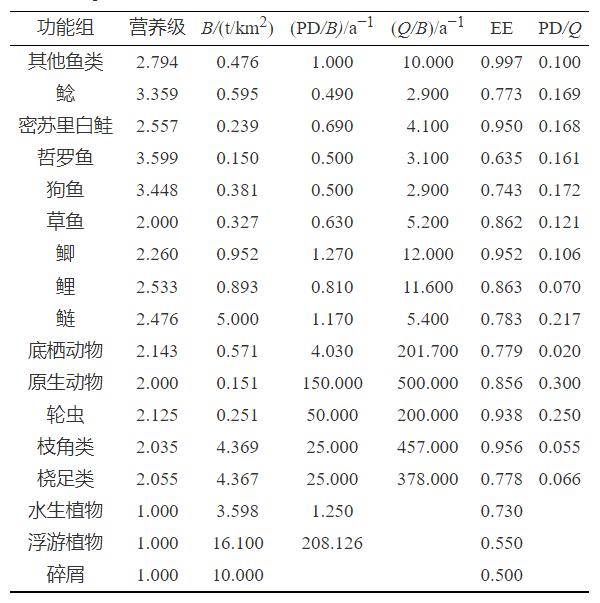
生物量(B)的获取。根据2014年开展的浮游生物调查工作,确定了浮游植物、桡足类、枝角类、轮虫、原生动物和底栖动物的生物量,鲢、鲤、鲫、草鱼、狗鱼、鲶和“其他鱼类”的生物量通过山口湖渔业公司的统计数据计算而得,水生植物和密苏里白鲑的生物量由Ecopath模型估计得出,哲罗鱼的生物量根据调研数据得出,结果见表1。
生产率(PD/B)的获取。水生植物和轮虫的PD/B参考宋兵[19]的研究,分别设为1.25和50a−1;桡足类和枝角类的PD/B参考Li等[20]的研究,均设为25a−1;原生动物的PD/B参考Kao等[21]的研究,设为150a−1;底栖动物参考李昌等[17]的研究,设为4.03a−1;除哲罗鱼的PD/B根据Ecopath模型中参数计算得出外,鱼类功能组的PD/B按照Fishbase网站(http://www.fishbase.org/)中设置的经验公式[22]获得,其中其他鱼类功能组中所含鱼类较多,按照经验公式计算每种鱼类的PD/B,乘以该种鱼类的生物量并求和,再除以其他鱼类功能组中每种鱼类生物量之和得到整体的PD/B,并根据模型的平衡情况进行调节。
消费率(Q/B)的获取。桡足类、枝角类和轮虫的Q/B参考宋兵[19]的研究,分别设为378、457和200a−1;原生动物的Q/B参考Kao等[21]的研究,设为500a−1;底栖动物的Q/B参考李昌等[17]的研究,设为201.7a−1。鲢、鲤、鲫、草鱼、狗鱼、哲罗鱼、密苏里白鲑和鲶的Q/B采用经验公式[22]进行估计,其他鱼类功能组根据包含的鱼种类结合经验公式进行估计。
生态营养效率(EE)的获取。浮游植物的EE参考Cremona等[18]的研究,设为0.55;山口湖水生植物的种类和分布较少,其主要捕食者草鱼的生物量在生态系统功能组中处于中间位置,参考类似区域[19]的研究结果,将水生植物的EE设为0.73。哲罗鱼处于食物链的顶层,其数量较少,参考Hossain等[23]的研究,将哲罗鱼的EE设为0.95,其他功能组的EE由Ecopath模型进行估计。
1.5 情景分析设置
为了探究山口湖生态系统中浮游植物的上行效应对高营养级生物的影响以及主要鱼类鲢对低营养级生物的影响,以2014年为基准年,使用Ecosim模型模拟浮游植物和鲢的变化。以2022年山口湖草鱼、鲤、鲫和鲢的生物量为起点,在浮游植物PD/B分别增加1%、5%、10%,降低1%、5%、10%情景下,预估浮游植物PD/B变化对山口湖草鱼、鲤、鲫和鲢的生物量的影响,不同情景依次命名为phy-s1、phy-s2、phy-s3、phy-s-1、phy-s-2和phy-s-3,对照组为control。搜索率通过物种的捕食能力及被其捕食的物种的生物量等参数进行计算,反映了物种对食物的获取能力,可以间接代表其生物量,以2022年山口湖浮游植物、桡足类、枝角类和轮虫生物量为起点,考虑在优势物种鲢搜索率分别增加1%、5%、10%情景下,预估鲢搜索率对浮游植物、桡足类、枝角类和轮虫的影响,不同的情景分别命名为silverC-s1、silverC-s2和silverC-s3,对照组为control。
为检验Ecosim模型的可信度,对Ecosim模型建模过程中需要用到的历史数据进行拟合,本研究中利用草鱼、鲢、鲤和鲫的2014—2021年历史渔业数值与Ecosim模型计算值进行比较,Ecosim模型计算值与利用捕捞量计算值的整体拟合趋势较好,平均离均差平方和为0.60,说明该模型的可信度较高[24-25],随着时间上新数据的出现可以继续对模型进行完善以使模型拟合效果更好[26]。
2. 结果与讨论
2.1 山口湖初级生产力影响因素分析
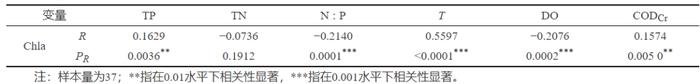
Spearman相关性分析见表2,由PR可知,Chla分别与TP、T和CODCr呈极显著正相关,相关系数(R)分别为0.1629、0.5597和0.1574,Chla分别与N∶P和DO呈极显著负相关,R分别为−0.2140和−0.2076。
表 2 环境因子对Chla影响Spearman相关性分析
Table 2. SpearmancorrelationanalysisofenvironmentalfactorsonChla
采用多元逐步回归模型对筛选到的4个指标(T、DO、TP、CODCr)与Chla建立响应关系模型:

TP是影响水体Chla的主要变量,影响程度显著高于其他3个指标,TP浓度的增加会显著提高Chla浓度,说明TP是影响山口湖初级生产力的主要因素。由藻类分子式(C106H263O110N16P)可知[27] ,氮和磷的原子比为16∶1,根据摩尔质量换算可知质量比为7∶1,即当N∶P大于7时,藻类生长受到磷含量限制。调查期间,山口湖TN浓度为0.681~1.771mg/L,TP浓度为0.022~0.080mg/L,年均N∶P约为53,表明山口湖是一个磷元素限制型的湖泊。因此,加强对山口湖磷输入的控制是降低生态系统初级生产力水平的关键。
2.2 山口湖的营养结构与能量流动特征
2.2.1 山口湖的营养结构分析
2014年山口湖Ecopath模型各功能组的输入与输出参数见表1,山口湖各功能组的营养级介于1.000~3.599,其中哲罗鱼的营养级最高,达到3.599。图2展示了山口湖中各功能组的食物网结构,山口湖食物网结构简单,17个功能组中11个功能组的营养级介于2~3,56%的功能组是杂食性功能组,杂食性鱼类有利于提高水层营养盐浓度,促进浮游植物生长,加速水体富营养化[28]。山口湖主要饲养鱼类鲢、鲤、鲫和草鱼的营养级分别为2.476、2.533、2.260和2.000,EE分别为0.783、0.863、0.952和0.862。其他鱼类功能组的EE为0.997,高于鲢功能组,但其他鱼类功能组的生物量显著低于鲢,说明下层营养级鱼类的生物量越大,上层营养级鱼类通过捕食作用所获得的能量越小,低能量传递效率使一部分能量累积,经由微生物作用后成为山口湖水体富营养化的能量来源[29-30]。
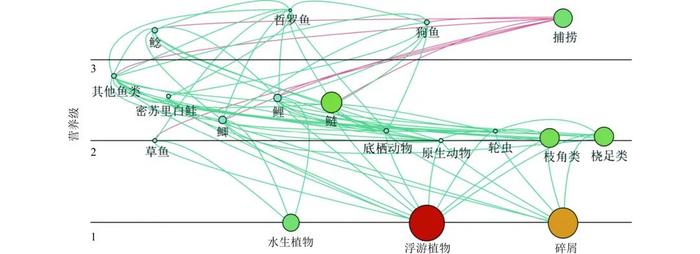
图 2 山口湖食物网结构
注:圆的面积代表B的大小,黑线代表营养级为1、2、3时的位置,绿线代表功能组间能量流动,红线代表捕捞。
Figure 2. FoodwebstructureinLakeShankou
2.2.2 山口湖生态系统的总体特征
根据Ecopath模型的输入参数进行计算,得到山口湖生态系统整体特征的指标(表3),其中系统总流量(TST),总初级生产量/总呼吸量(TPP/TR)、连接指数(CI)、系统杂食性指数(SOI)以及Pedigree指数是描述生态系统总体状况的常用变量。
表 3 山口湖生态系统指标统计
Table 3. EcosystemindexesstatisticsofLakeShankou
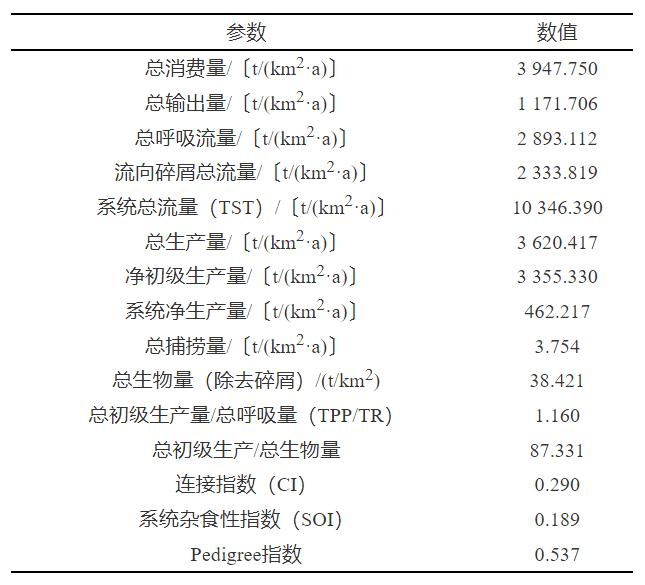
TST是生态系统中所有功能组能量流动的汇总。气候条件是影响湖泊TST的重要因素。山口湖的TST为10346.390t/(km2·a),比高海拔湖泊海格湖〔39685t/(km2·a)〕[31]低,比纬度更高的瑟峡湾〔6525t/(km2·a)〕[32]高。海格湖位于埃塞俄比亚的阿姆哈拉州,受热带季风气候影响,比山口湖更适宜水生生物的生存,而瑟峡湾位于北纬60°附近的挪威西部卑尔根地区,受亚寒带针叶林气候影响比山口湖更严寒。结合相关性分析可知,在山口湖中T和Chla呈正相关,T越高,Chla越高,TST相应增高。
生态系统的成熟性和稳定性是反映其健康程度的重要因子[33],TPP/TR越接近1则表明生态系统的成熟性越高[34]。山口湖TPP/TR为1.16,相较于太湖(3.85)[35]、千岛湖(3.73)[8]、白洋淀(2.58)[11]等表现出更高的成熟性,这说明山口湖生态系统的自我调节能力较强,受人类活动影响较小,尤其是每年长达半年的冰封期隔绝了外源污染物的流入,藻类成为主要的初级生产者,营养物质得到有效循环和保持。CI反映了系统生物之间彼此联系的复杂程度[36],CI和SOI均与生态系统成熟度相关[30],山口湖的CI和SOI分别为0.290和0.189,相较太湖(0.206和0.042)[35]、千岛湖(0.222和0.087)[8]、白洋淀(0.216和0.044)[11]等表现出更高的成熟性和稳定性。作为高寒山地湖泊,其水生生态系统的生物之间的相互作用较强,能更好地预防冰封期带来的初级生产者生产率下降的问题。
Pedigree指数可以检验模型参数来源的可靠性,Colléter等[37]对Ecobase数据库中433个Ecopath模型进行汇总,分析提供Pedigree指数的34个模型,发现Pedigree指数介于0.137~0.743,平均值为0.472,对Ecopath模型在山口湖生态系统应用的可靠性进行检验,其Pedigree指数为0.537,说明山口湖参数来源的可靠性较高。
2.2.3 山口湖能量流动分析
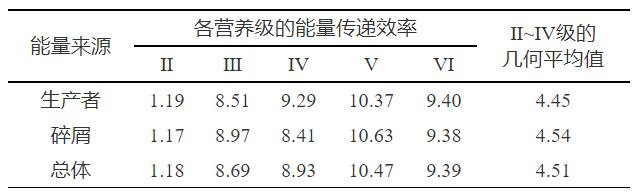
各功能组在食物网的不同食物链中有不同的整数营养级,为了探究山口湖中整数营养级的能量传递效率,使用Ecopath模型进行汇总计算,表4展示了山口湖Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ营养级的能量传递效率,可知来自碎屑和来自生产者的能量传递效率接近,而营养级Ⅳ来自生产者的传递效率明显偏大,因为山口湖中的能量来源主要是生产者,占生态系统能量来源的60%。从总体来看,营养级Ⅱ的传递效率较低,为1.18%;营养级Ⅲ、Ⅳ、Ⅴ和Ⅵ的传递效率分别为8.69%、8.93%、10.47%和9.39%。低营养级的传递效率低导致很多能量不能在生态系统中正常循环消耗,积累在水体中会导致湖泊生态系统恶化。山口湖的能量流动主要集中在Ⅱ级以上,原因是Ⅱ~Ⅲ级的鱼类主要以轮虫、枝角类以及桡足类为食,肉食性鱼类如哲罗鱼等以营养级较低的鱼类为食。山口湖和白洋淀的传递效率趋势相似,结合Zeng等[11]对白洋淀的分析以及山口湖的实际情况,发现山口湖大部分的初级生产力没有被利用,这与鲢、鳙等滤食性鱼类资源被过度开发有关。冰封期长也是导致山口湖生态系统能量传递效率偏低的重要原因,冰封期阶段水生生物生长代谢速率慢,不同营养级之间的能量传递效率也随之降低。
表 4 山口湖能量传递效率统计
Table 4. StatisticsofenergytransferefficiencyinLakeShankou %
2.2.4 功能组间相互作用及关键种分析
图3展示了山口湖生态系统各功能组的相互影响。由图3可以看出,作为山口湖的初级生产者,浮游植物和水生植物对上层生物的生长具有正向促进作用。草鱼对水生植物的捕食作用占据了草鱼能量向营养级上方传递的大部分,水生植物对草鱼有很明显的正向影响,山口湖中主要的经济鱼类鲢对水产品捕捞也有明显的正向贡献。其他鱼类功能组对哲罗鱼有明显的负向影响,狗鱼对密苏里白鲑有明显的负向影响,因为对鲢的大量捕捞减少了肉食性鱼类的食物来源,捕捞作用对山口湖中的鱼类尤其是肉食性鱼类鲶和狗鱼的生长有负向作用。对不同功能组间的相互作用分析发现,浮游植物对山口湖几种主要鱼类(鲫、鲤、鲢)有着明显的正向影响,这为设置情景分析提供了参考依据。
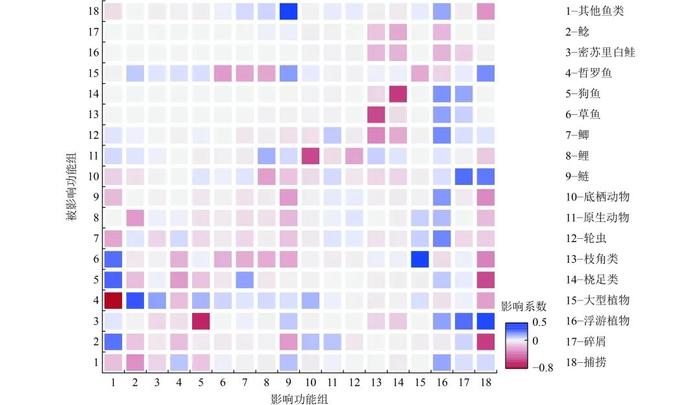
Figure 3. MixedtrophicimpactdiagraminLakeShankou
由各功能组的关键种指数(图4)可知,山口湖生态系统的关键种是其他鱼类功能组。关键种指的是对于维持生态系统起重要作用的物种[38],其他鱼类功能组的生物量不高但EE最高,说明其对维持山口湖生态系统群落的组成和多样性具有决定性作用。狗鱼、底栖动物和鲶功能组的关键种指数低于其他鱼类功能组,高于鲢的关键种指数。在渔业管理的过程中除了关注鲢等主要鱼类,还要注重对山口湖鱼类生物多样性的保护[39],这是山口湖可持续健康发展的关键。
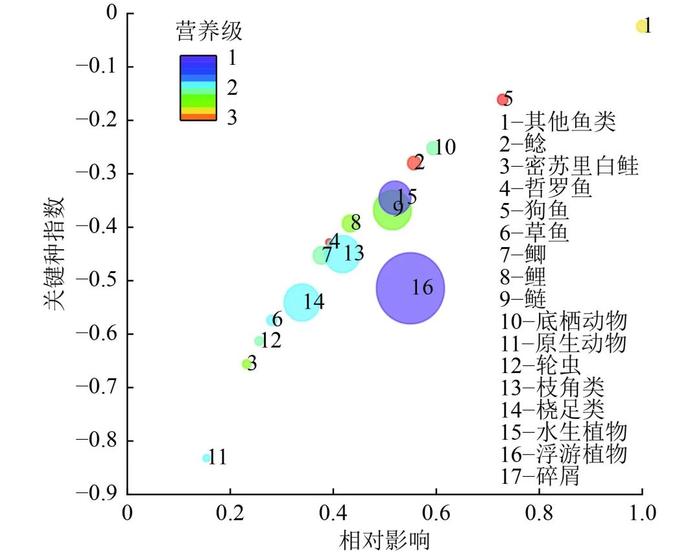
注:圆的大小和颜色分别代表每个功能组生物量和营养级。
Figure 4. KeystonespeciesindexinLakeShankou
2.3 山口湖生态系统中上行、下行效应的探究
2.3.1 浮游植物的上行效应情景分析
对照组中,草鱼、鲫、鲤和鲢4种鱼类的相对生物量2014—2017年增至极大值(1.28、1.21、1.25和1.30),2017—2019年降至极小值(0.54、0.60、0.58和0.51),2019年之后呈现不同的变化趋势(图5)。草鱼的相对生物量2019—2026年继续增加,达到极大值(2.37)后下降;鲫的相对生物量2019—2022年继续增至极大值(1.62),随后降至1.00左右;鲤的相对生物量2019—2024年继续增加到极大值(1.94)后降至1.10左右;鲢的相对生物量2019—2023年继续增加到极大值(1.62)后降至1.40左右。
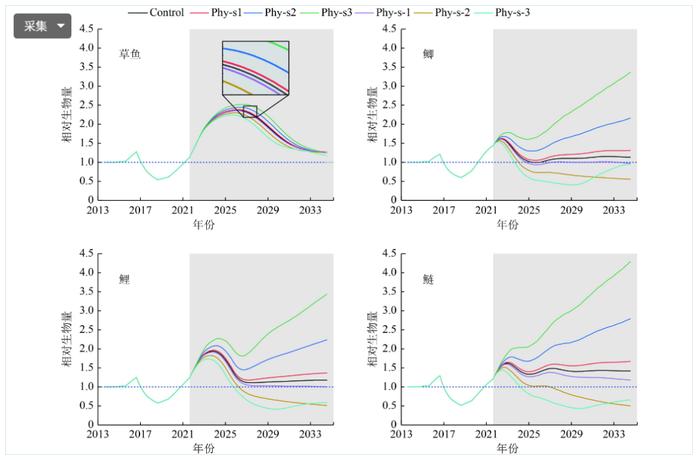
Figure 5. Bottom-upeffectsofphytoplanktononmainfishinLakeShankou
与对照组相比,草鱼在藻类增加和减少的6种情景下变化不大,鲫、鲤和鲢的相对生物量会随着藻类PD/B的增加而增加,其中鲢的相对生物量变化最明显,如果藻类的PD/B每年增加10%,到2034年末鲢生物量可以达到2014年的4.28倍。藻类PD/B变化对鱼类的影响呈现鲢>鲫、鲤>草鱼的趋势,取食方式以及食物组成是产生差别的主要原因。鲢是选择性弱的滤食性鱼类,对藻类生物量变化敏感,鲫和鲤属于捕食性鱼类,草鱼以水生植物为食。对于鲢、鲤和鲫而言,浮游植物10%减少的情景并不会比5%减少的情景对鱼类生物量的影响更加明显,反而在2032年末会有回增,原因是藻类并不是鱼类的唯一食物,藻类PD/B下降到一定程度会导致浮游动物对水体中有机碎屑的利用增强,促进浮游动物生物量的增加,导致鱼类生物量的回升。总之,藻类PD/B增加会引起鱼类生物量的增加,藻类不仅可以直接作为鱼类的食物来源,还通过促进浮游动物的生长间接为鱼类提供更多的食物。
2.3.2 鲢的下行效应情景分析
2014—2021年对照组中浮游植物、桡足类、枝角类以及轮虫的相对生物量保持在1附近(图6)。从年际变化上看,浮游植物相对生物量呈现先增加后下降再趋于稳定的趋势,桡足类、枝角类及轮虫相对生物量呈现相反的趋势,即先下降后增加再趋于稳定,这说明山口湖生态系统中鲢优先以尺寸较大的藻类和浮游动物为食,粒径较小的藻类如蓝藻等很容易被过滤掉,因而控藻效果不明显。与对照组相比,从2022年开始鲢搜索率每年分别增加1%、5%和10%,浮游植物的相对生物量呈现增加趋势,桡足类、枝角类及轮虫相对生物量呈现减少的趋势,进一步验证了鲢以浮游动物为主的饮食结构。在鲢搜索率增加10%的情境下,与对照组相比桡足类、枝角类和轮虫的相对生物量分别下降了0.33%、2.17%和4.50%,说明鲢对轮虫的摄食能力更强。
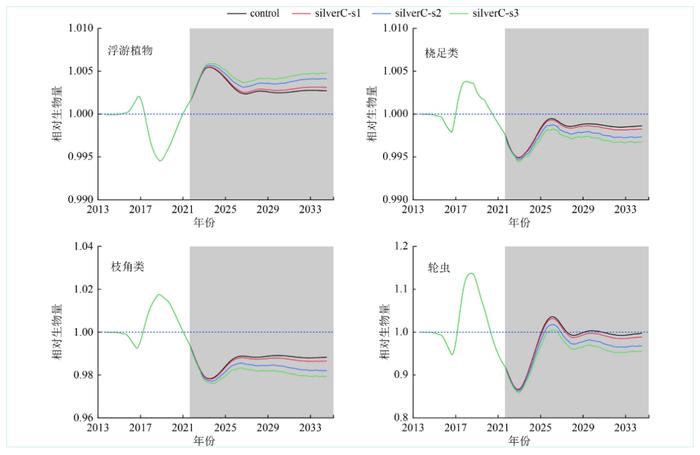
图 6 山口湖中鲢对浮游生物的下行效应
Figure 6. Up-downeffectsofsilvercarponplanktoninLakeShankou
结合对鲢食性的研究发现[40-41],鲢更易消化超过30μm的浮游生物,而调查数据显示2014年山口湖以蓝藻、绿藻和单细胞硅藻(菱形藻属、平板藻属、小环藻属等)为主,部分单细胞硅藻的大小不到10μm,鲢不能很好地消化利用,甲藻、隐藻等非优势藻类和浮游动物是利于鲢消化利用的主要食物来源。Yin等[42]发现洱海的日本胡瓜鱼被大量捕捞时,浮游动物的生物量上升。浮游植物是枝角类、桡足类以及轮虫等浮游动物和鲢共同的重要食物来源,当浮游动物对浮游植物的影响大于鲢对浮游植物的影响时,引起浮游植物相对生物量随时间的延长略微下降,2027年后鲢搜索率的增加提高了其对浮游动物的捕食能力,浮游动物对浮游植物的影响减弱,浮游植物的相对生物量开始增加。这说明对于高寒山地湖泊生态系统而言,鲢控藻的效果并不明显,结合山口湖初级生产力影响因素分析可知应加强对水体营养盐尤其是对TP的控制,才能达到较好的控藻效果。
3. 结论
(1)水温、溶解氧和总磷是影响东北山地山口湖初级生产力的主要因素,影响程度表现为总磷>水温>溶解氧,除溶解氧外,其他因素均对Chla含量产生正向影响。
(2)山口湖是一个成熟的生态系统,营养级Ⅱ的传递效率仅为1.18%,能量流动集中在Ⅱ~Ⅳ级,浮游植物对鲫、鲤、鲢等鱼类呈现明显的正向影响。山口湖生态系统的关键种为其他鱼类功能组,如鳌花、鳊花等低生物量鱼类,维持山口湖生物多样性有利于保持生态系统的平衡。
(3)浮游植物PD/B在0%~10%变化时,对山口湖中鲫、鲤和鲢的相对生物量有明显的促进作用,对草鱼相对生物量的影响较小;浮游植物PD/B在−5%~0%变化时,鲫、鲤和鲢的相对生物量显著降低;PD/B在−10%~−5%变化时对鱼类相对生物量的影响较小。说明浮游植物PD/B持续减少会使鱼类找到替代性的食物来源。
(4)随着鲢的搜索率增加,山口湖生态系统中浮游植物和桡足类、枝角类、轮虫等浮游动物的相对生物量呈现相反的年际变化趋势,浮游动物对浮游植物的影响显著,浮游植物的相对生物量逐渐增大,浮游动物的相对生物量逐渐减小,进一步说明了鲢以浮游动物为主的食物组成。对于东北山地湖泊山口湖而言,鲢控藻的效果不好,应加强对水体营养盐尤其是总磷的控制以减少藻类繁殖。
向上滑动阅览
参考文献



